
私たちの環境において対象物がどこにあるのかを知るために、私たちが行わなければならない最初のことは、対象物を相互に分離し、そして背景から分離しなければならない。それから知覚機構は、私たちからの距離と対象物の位置を決定できる。
対象物の分離
私たちの網膜に写し出される像は、変化する明るさと色の寄せ集めである。私たちの知覚機能は、その寄せ集めをどうにかして背景に投影された一組のまとまりのある対象物に体制化する。この種の体制化はゲシュタルト心理学にとって大きな関心ごとであった(ゲシュタルト心理学は20世紀の初めにドイツで始まった心理学の一つの方法)。
ゲシュタルト心理学は対象物全体あるいは形全体を知覚することの重要性を強調した。そして私たちがどのように、対象物を体制化するのかを説明するために多くの原理を提案した。
図と地
知覚の体制化の最も初歩的な形は、二つ以上の異なる領域を持つ刺激において、私たちは通常、刺激のある部分を図として、そして残りを地(あるいは背景)としてみる。ということである。図としてみられた領域は興味のある対象物を含んでいる。図は地よりも立体に見えるし、地の前にあるように見える。
図Aのa)は、図-地の体制化が多義的であることを示している。
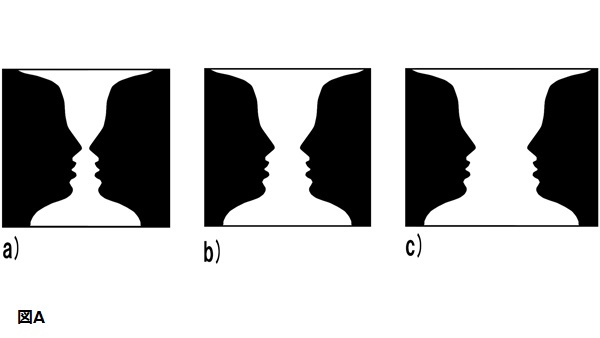
あなたがこの図柄を見るとき、互いにじっと見つめ合っている横顔の影に見えるかもしれないし、装飾用の壺に見えるかもしれない。壺は黒い地に対して、白に見えるが、顔は白い地に対して黒である。貴方がしばらくの間、b)を見ていると、二つの図柄の体制化が意識の中で交互に起こることに気づくだろう。
それは図と地の体制化が刺激ではなく、あなたの心の中にあることを示している。また、顔と壺が決して一緒には現れないことに気づくであろう。あなたは両方が可能であることを「知っている」が、同時に両方を「見る」ことができない。
一般的に言えば、領域や形が小さいほど、それは図として見られやすい。このことは、a)、b)、c)を比較することによって示される。白の領域が狭いほど壺に見えやすく、黒の領域が狭いほど顔に見えやすい(Weisstein&Wong,1986)。これらの図-地の原理は単純な刺激に限定されない。
視覚は図-地関係の最もな顕著な拠り所であるけれども、私たちは他の感覚においても図-地関係を知覚できることに注目すべきである。たとえば、私たちは戸外の騒音を背景にして鳥のさえずりを聞くかもしれないし、バイオリン以外のオーケストラの音楽を背景にして聞くかもしれない。
対象物の群化
私たちは単に地を背景にした対象物を見ているだけでなく、同様に特定の群化している対処物も見ている。点の単純な模様でさえ、私たちがそれらを見ているときには群になる。これを説明するために図1のa)に示された行列をまずは見てみよう。
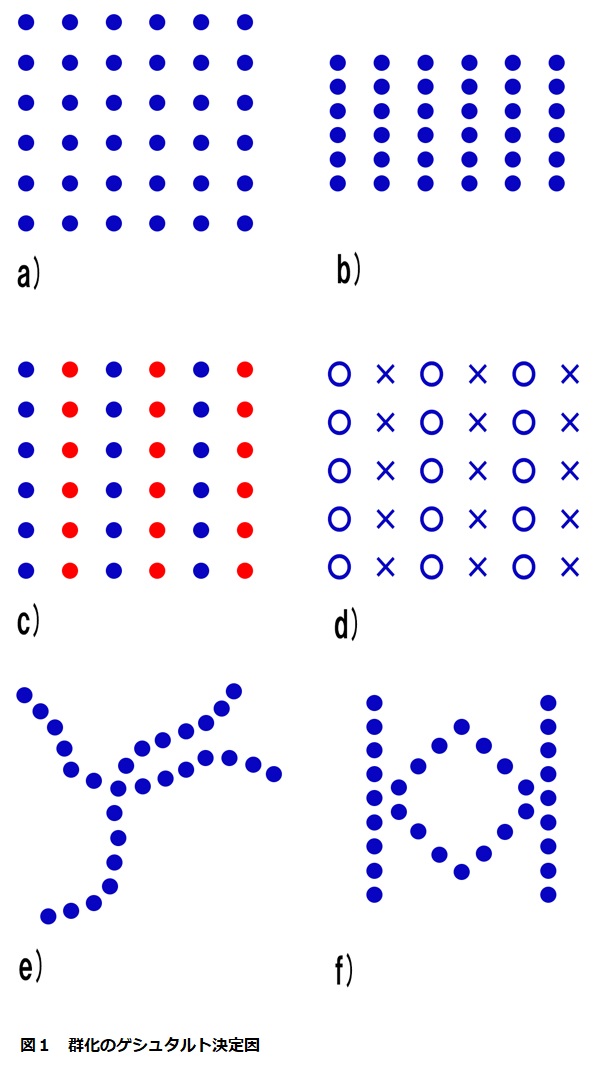
これらの点は上下に等しい間隔で配列されている。よってそれらは行あるいは列に体制化されているように、あるいは斜めの小道に沿って置かれているようにさえ見える。したがって、これは図Aで説明されたのと同じ原理に従う多義的模様である。一度に一つの体制化だけが見られ、そしてときどきこの体制化が別のものに無意識のうちに変わるだろう。
ゲシュタルト心理学者はこの種の点の模様に関する群化の多くの決定因を提案した。たとえば、もし点の垂直の間隔が減少すれば、図1のb)のように、縦の行はほとんど列のように見えるだろう。これは近接による群化である。点の間隔を変える代わりに要素の色や形を変えるのならば、私たちは類同に基づいて点を体制化することができる(図Aのc),d))。
もし私たちが点を動かして、点でできた2本のの交差する破線を作ろうとするなら、よい連続によって群化するだろう(図Aのe))。そして私たちが点線を使って空間を囲むのならば、閉合による群化を見ることになるだろう。
この最後の例では、例えのその模様がお互いの上にきちんと積み重ねられた二つのよく知られた文字(Mの上のW)であったとしても、私たちは互いに向かい合っている文字(KとKの鏡像)であったとしても、私たちは2本の垂直線の間に置かれたひし形を見ることに注目しよう。これはゲシュタルトの群化の決定因が説得力のある性質を持っていることを物語っている。これらの決定因は与えられた模様の範囲内で可能な最も安定した、矛盾のない、そして単純な形を創造するのに役立っている。
視覚的な群化についての現代の研究は、ゲシュタルト決定因が知覚に強い影響力を持っていることを示している。たとえば、一連の研究において、近接に基づいたより大きな視覚的群化の部分になっている視覚的標的は、その群の外側にあるように見える同じ目標よりも検出するのが難しかった(Banks&Prinzmetal,1976;Prinzmetal,1981)。
別の一連の研究において、色と形において非標的に似ていない目標は、より似ている標的よりも検出が容易であった(Triesman,1986)。さまざまな非標的間の類同でさえ重要な影響を持っている。すなわち日標的の類同が増すほど、標的は検出しやすくなる。
それは標的が背景と異なった図として「ポンと現れる」からである(Duncan&Humphreys,1989)。最後に、ゲシュタルトの決定因と結びついた確かな錯覚がある。人は異なる群の要素間の距離よりも、知覚群内の要素間の距離の方を短く判断する(Coren&Girgus,1980;Enns&Girgus,1985)。これらの結果のすべては、私たちが視覚経験を体制化するという点で、視覚的群化が大きな役割を果たしているということを示している。
知覚的群化は主に視覚において研究されてきたが、群化に関して同様の決定因が聴覚においても現れる。これに関する多くの証明は、楽音知覚を研究する研究者たちによってなされている。近接は聴覚において明らかに作用する。たとえば第2打と第3打の間に休止が入る、ドラムを四つたたく音は二つの対として聞こえるだろう。
同様に、時間的に接近している音符のひとまとまりは、群化されるだろう(ベートーベンの第5交響曲のDUH-DUH-DUMMMという導入部におけるように)。音の高さにおいて近接した音符もまた群化されるであろう。音楽はしばしば、二つの旋律が同時に生じる、対位旋律を含んでいる。
聞き手は、注意を向けた旋律が図、注意を向けない旋律が地になるように、旋律間で注意を移すことができる。これはしばしば、二つの旋律が異なったオクターブなので、可能である。そのため、一つの旋律内の音符はお互いに接近しているが、旋律間の音符は接近していない。類同と閉合もまた、音の高さや、より複雑な刺激を聞くのに重要な役割を果たしていることが知られている(Bregman,1990)。
距離知覚
ある対象物がどこにあるのかを知るために、私たちはその距離や奥行を知らなければならない。対象物の奥行を知覚するすることは努力を要しないように思われるが、私たちは奥行の次元に直接接近できないので、それは実際には驚くべき功績である。
それによって、私たちが先に議論した、多対一問題の一つの形に至ることになる。網膜は、三次元世界が映し出される二次元の面である。それゆえ、網膜は直接に高さと幅を移すが奥行の情報は失われ、結果として奥行手掛かり(depth cues)として共通に知られている情報の難解な要素に基づいて、何とか再構成されなければならない。
奥行手掛かりとは、ある対象物の奥行についての情報を、論理的あるいは数学的に提供するさまざまな種類の視覚情報である。奥行手がかりは両眼手がかりと単眼手がかりに分類される。
両眼手がかり
なぜ私たちやほかの動物たちは、一つではなく二つの眼をもっているのか。それには二つの理由がある。いくつかの動物、たとえばサカナは頭ののどちらかの側に眼を持ち、頭や身体を動かさないで、周りに大部分の世界を見ることができる。
ほかの動物、たとえば人間に関しては頭の前に二つの眼を持ち、両方の眼は同じ方向を向いている。人間はある瞬間にサカナ以上には世界を見ることができないが、奥行を知覚するために二つの眼を使うことができる(一方の眼を覆ったままでは、対象物にどれだけ接近しているのかについて、通常よりもほとんど感覚がない)。
二つの眼で見ることは奥行を知覚する上で有利である。なぜなら、両眼は頭で分離し、それはそれぞれの眼が同じ場面についてわずかに異なった見え方をすることを意味するからである。あなたは、右手の人差し指を顔に近づけ、まず一方の眼だけを開けて指を見て、次にもう一つの眼だけを開けて指を見ることによって、これを容易に説明できる。
両眼差異(binocular disparity)という用語は、それぞれの眼による見え方の差異を指すために使われる。視差は近い範囲で見られる対象物に関して最も大きくなり、対象物が遠ざかるほど小さくなる。3~4メートル位以上になると、それぞれでの眼で見える見え方の差異は非常に小さくなるので、両眼差異は奥行に関する手がかりとしての効果を失う。しかし対象物に手を伸ばしたり、障害物の周りを通過するというような多くの日常的な課題では、それぞれの眼で見える見え方の差異は奥行についての有力な手がかりになる。
両眼視力を持つ人間やほかの動物では、脳の視覚の部分は、ある対象物の二つの視覚像が比較されたときに、どのくらい離れているかによって、空間の様々な位置に対象物を割り当てるため両眼視差を使っている。もしある対象物の視覚像が二つの見え方で同じ場所にあるならは、脳は、これを両眼の眼が固定している位置であると仮定する。もしあなたの顔に指を近づけたときの二つの見え方のように視覚像の差異が大きいならば、脳は対象物がより近くにあると結論する。
両眼視差は日常的な世界において私たちに奥行を知らせることに加え、実際には何もないときに奥行が見えるように目をだますためにも使われる。これを行う一つの方法は、実体鏡と呼ばれる装置を使うことである。実体鏡は、それぞれの眼にわずかに異なった写真を見せる。この両眼視差と同じ原理のおもちゃや、色眼鏡や偏光フィルターのメガネを装着し「特殊効果」を持つ立体映画でも使われている。それは選択的に、一方の眼に一つの像、他方の眼にそれとはわずかに異なった像が到達するようにできている。
単眼手がかり
指摘したように、両眼手がかりは比較的近くにある対象物に限定して使われる。遠い雲、都市の風景、山のような遠く離れたところにある対象物についてはどうであろうか。この点では、両眼手がかりは比較的役に立たず、単眼手がかりとして知られる他の手がかりを使わなければならない。
このように、視覚機構の課題は簡単ではない。本質的に、その機構は結論に達するために、環境において利用できる情報を寄せ集めて利用しなければならない。それは、刑事が殺人者はだれかを判断するために、殺人者について利用できる証拠をすべて使わなければならないのと同様である。
図2は五つの単眼手がかりを説明している。それに一つ加えたものが以下である。
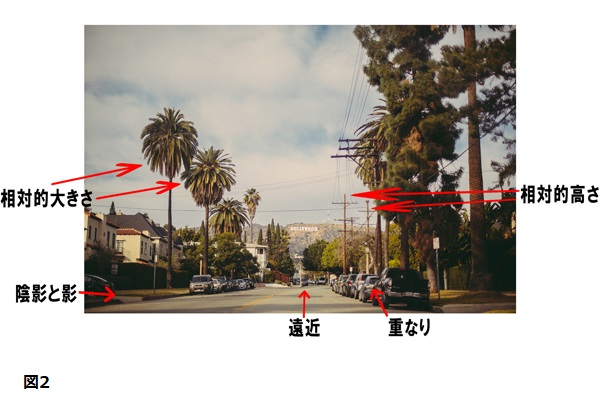
1)相対的大きさ。もしある視覚像が大きさだけが異なる同様の対象物の配列を含んでいるならば、見る人はより小さい対象物を遠くに離れているものとして解釈する。
2)重なり。もし一つの対象物が他の対象物の視界を妨げるように位置しているならば、見る人は重なってる対象物をより近くにあるものとして知覚する。
3)相対的高さ。同様に対象物の中で、地平線により接近して見えるものは、遠くに離れているものとして知覚される。
4)陰影と影。ある場面で表面が直接の光を受けていることを妨げられているときはいつでも、影が落とされている。、もしその影が光をさえぎっている同じ対象物の一部に落ちているならば、それは付着した影あるいは単に陰影と呼ばれる。もしそれが影を落としている対象物に属さない、ほかの表面に落ちているならば、それは投影した影と呼ばれる。両方の種類の影は、対象物の形、対象物間の距離、そして場面における光源の位置に関する情報を私たちに与えてくれる。その場面の奥行を知る重要な手掛かりである(Coren,Ward,&Enns,1999)。
5)遠近。ある場面での平行線がその視覚像において縮まるように見えるとき、平行線は遠方で消失するものとして知覚される。
6)運動。あなたが素早く動いてると(高速の列車に乗っているときなど)近くの対象物は反対方向へすばやく動き、遠い対象物ほど(なお反対方向であるけれど)ゆっくり動くようだということに、今まで気づいたことがあるだろう。月のように離れた対象物は全く動いているように見えない。動いているように見えるこれらの対象物の速度差は、私たちの距離を知るのに手がかりを提供してくれる。それは運動視差と呼ばれる。
運動知覚
最後の単眼手掛かりである運動は、定位に関係する次の重要な話題を私たちに提供してくれる。環境の中で効果的に動き回るためには、私たちは静止した対象物の位置だけでなく、動いている対象物の道筋も知る必要がある。たとえば、一区画離れたところから私たちの方へ向かってくる車は、私たちが道路を横断し終わるまでは交差点に達しないだろうというように、私たちは運動を知覚しなければならない。
ストロボスコープ運動
何がわたしたちに運動を知覚させるのであろうか。もっとも単純な考え方は、対象物の視覚像が網膜を横切って動くときは私たちはいつでもその対象物が動いていると知覚するということである。しかし、私たちは、網膜上で何も動いていないときでさえ、運動を見ることができるところを見ると、この答えはあまりに単純すぎることがわかる。
図4に示されているこの現象は1912年ウェルトハイマー(Wertheimer)によるストロボスコープ運動の研究によって証明された。ストロボスコープ運動はもっとも単純には、暗黒の中で光を発し、そしてそれから2,3ミリ秒後に最初の光の位置の近くでもう一つの光を発することによって作り出される。その光は、実際の運動とは区別できないという点で、一方の場所から他方へ動くように見える。a)における静止したコマの連続は、b)に示した知覚内容になる。
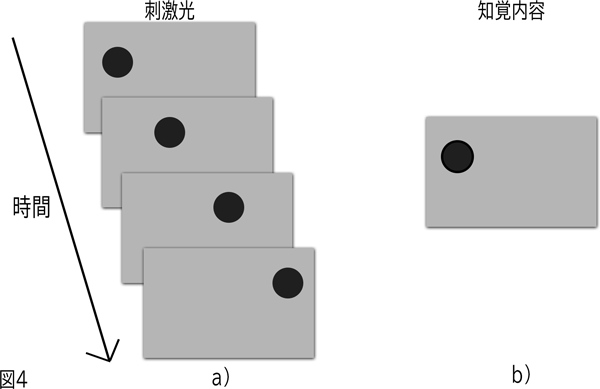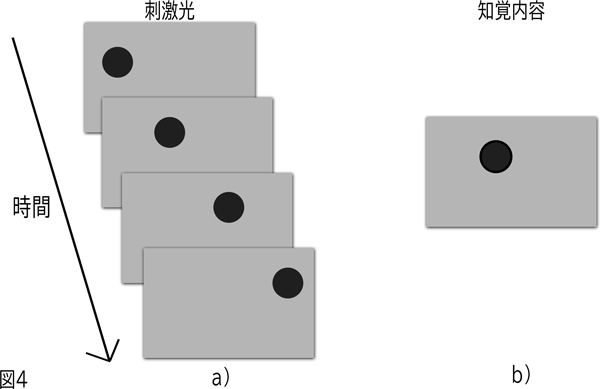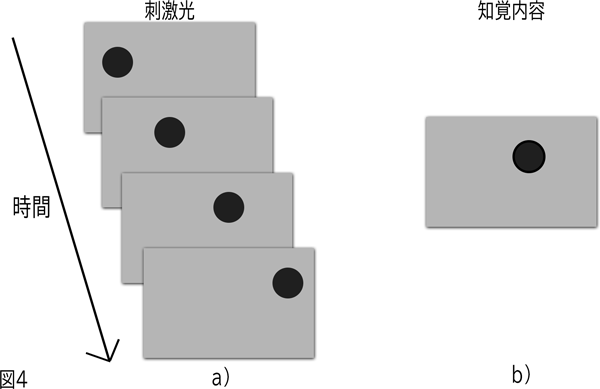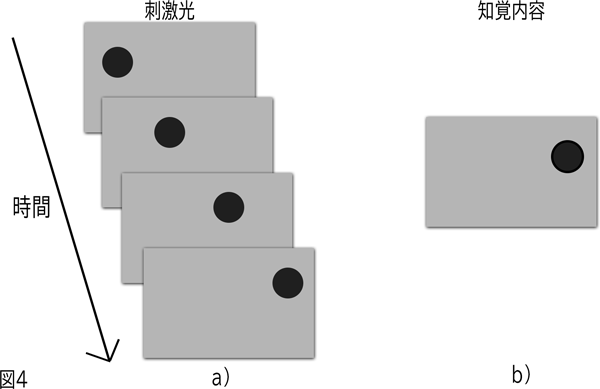
ストロボスコープ運動に関するウェルトハイマーの証明は、たんなる学術的な実演にとどまらない。その現象は多くの現代の画像技術にとって欠くことができないものである。主要な例は映画であり、映画の中で私たちが知覚する運動がストロボスコープ運動(stroboscopic motion)である。多くの人が知っているように、映画は単に一連の静止した写真(すなわち「コマ」)であり、それぞれの写真は先行するものとわずかに異なっている。このようにコマはスクリーンに連続して表示される。
実際運動
わたしたちの視覚系は実際運動である。すなわち、空間におけるすべての中間点を通る対象物の動きにも敏感である。しかし、日常の条件下でそのような運動を分析することは、驚くほど複雑である。網膜上のいくつかの運動経路は、(私たちが読書をしているときに生じるような)静止した場面上の眼の動きによるものと考えなければならない。
その他の運動経路は、(トリが私たちの視野に入ってくるように)動いている対象物によるものと考えなければならない。さらに、網膜像が静止しているいくつかの対象物は、(私たちが飛んでいるトリを眼で追っているときのように)動いているように見えなければならない。一方、網膜像が動いているいくつかの対象物は、(私たちの眼は飛んでいるトリを追っているので、静止した背景が網膜を横切る運動をたどるときのように)静止しているように見えなければならない。
それゆえ、運動に関する私たちの分析がかなり相対的なものであるということは、驚くべきことではない。私たちは、背景が一定の色で、動いている対象物だけが見える(相対的運動)時の動きをよりよく検出する。ある相対的な動きの様式は、三次元対象物の特定の形にとって有力な手がかりとして役に立つことさえある。
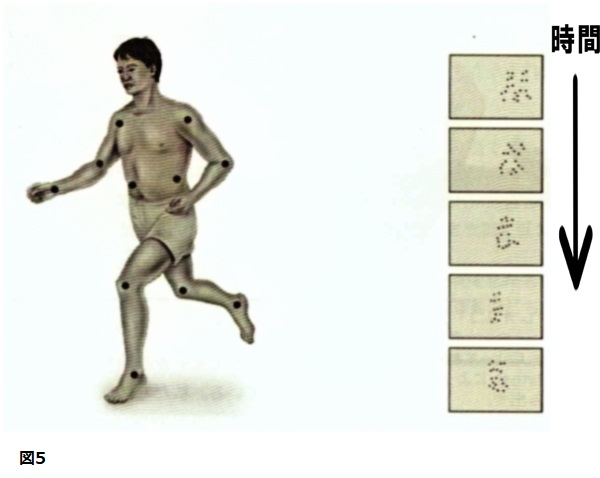
たとえば、研究者たちは図5で説明されている運動表示器が、たとえお互いに相対的に動く、12個(もっと少ないことさえある)の光点からなるものとしても、参加者は人間の活動図であると容易に確認させ得ることを見出している(Johansson,von Hofsten,&Jansson,1980)。これらの表示器を使っている他の研究では、参加者は彼らの友人を確認でき、足首についた光だけで、そのモデルが男性か女性かを言うことさえできた(Cutting,1986)。
実際運動の研究において、ほかの重要な現像は選択的順応(selective adaptation)である。これは、私たちが見続けている運動や類似の運動に対する感度を低下させるが、方向や速度において有意に異なっている運動に対してそうではない、という点で順応は選択的である。
たとえば、もし上向きに動いている縞が見えるならば、上向きの運動に対する感度は低下するが、下向きの運動を見る能力は影響されない(Sekuler,1975)。ほかの種類の順応のように、通常私たちは感度の損失に気づかないが、順応によって生じる残効には気づいている。もし数分間滝を見続け、それからそのそばの断崖を見ると、その断崖は上向きに動いているように見えるだろう。ほとんどの運動はそのような残効を常に元の運動とは反対方向に引き起こすであろう。
脳は実際運動の近くをどのように実行しているのだろうか。実際運動のいくつかの側面は、視覚野の特定の細胞によって符号化されている。これらの細胞はある運動には反応し、ある運動には反応しない。そして、各細胞は運動の一つの方向と速度に最もよく反応する。
そのような細胞の存在に関する研究からもたらされている。その研究では、動物がさまざまな運動刺激を見ている間、動物の視覚野における単一の細胞の反応を実験者は見ている。そのような単一の細胞の記録をする研究は、特定方向の運動に同調する皮質細胞を見いだした。頭の方へ向かって動く対象物を検出するのにとくに同調する細胞も存在する。それは生き残るために明らかに必要な能力である。(Regan,Beverley,&Cynader,1979)。
これらの分化した運動細胞が、選択医的順応の説明になり得る。おそらく、たとえば上向きの運動に対する選択的順応は、上向きの運動を専門に扱っている皮質細胞が疲労するために生じる。下向きの運動を専門に扱っている細胞はいつものように機能しているので、それらが処理を支配し、そしてその結果下向きの運動の残効を生じることになる。
しかし実際運動の神経的基礎は、特定の細胞の賦活だけでもたらされたものではない。私たちは(夜の飛行機のような)暗い中を動いている光を発する対象物を追うとき、運動が見えている。私たちの眼は対象物の後を追うので、その像は網膜上でほとんど動かない。
それにもかかわらず、滑らかな連続した運動を知覚する。それはなぜだろうか。私たちの眼がどのように動いているかについての情報は、脳の前部の運動領域から視覚野へ送られ、私たちが見る動きに影響を及ぼす、というのがその答えだとであるかのように思われる。
本質的に、運動機構は、網膜上の通常の働きが欠如しているのは視覚機構のせいであるという情報を伝えており、視覚機構はそれからすぐその欠如を修正する。もっと多くの普通に見える状況では、眼球運動と網膜像の大きい動きの両方がある。
視覚機構は知覚した動きを決定するために、これらの二つの情報源を結合させなければならない。あなたはまぶたで眼球を優しく押し上げることによって、この調整の結果を例証できる。世界は動いているように注目しよう。これは、世界があなたの網膜を横切って動いているためである。しかし、運動領域からの通常の信号はない。脳がそれを解釈できる唯一の方法は、世界それ自体が動いているかどうかである。

コメント