
ニューロンは神経系の基本単位であり、情報を他のニューロンや内分泌線、筋肉に伝える細胞で、形状や大きさは様々であるが、一定の共通の特徴をもっている。
ニューロン(neuron)の構造
細胞体(体)からは樹状突起(dendrites)と呼ばれる多くの短い突起が出ていて、隣接する細胞から神経インパルスを受け取り、軸索(axon)は細胞から伸びている細い管で、他のニューロン(もしくは、筋肉、内分泌腺)に情報を伝える働きをする。
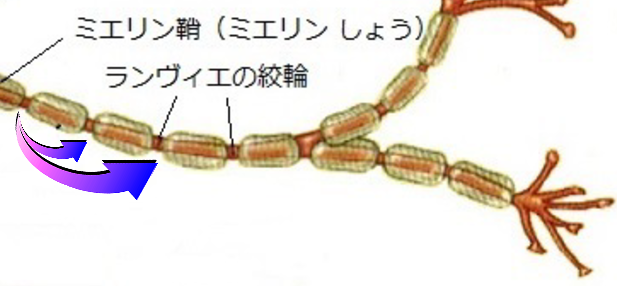
また、軸索には分岐しているものもあり、この様な分岐を側枝と呼ばれ、終端ではたくさんの枝に別れ、シナプス終末と呼ばれる小さな膨らみの終末ボタン(termina buttons)までになる。軸索はミエリン鞘((depolarized)あるいは髄鞘)と呼ばれる電気絶縁体で覆われていて、1~3mmの間隔で覆われていない部分がある。
この部分をランビエ絞輪(nodes of ranvier)と言い、この部分は軸索が細胞外液にさらされているため電気抵抗が低くインパルスの伝導速度が増す。
終末ボタンは、実際には隣接するニューロンには密着していなく、受けて側のニューロンの細胞体や樹状突起との間には、わずかな隙間があり、この接合部位はシナプスと呼ばれ、隙間そのものはシナプス間隙(synaptic gap)と呼ばれる。
神経インパルスが、軸索を伝わって終末ボタンに到達すると、神経伝達物質がシナプス間隙に放出され、次のニューロンを刺激する化学物質の分泌を引き起こす。それによってインパルスを1つのニューロンから次のニューロンへと伝える。
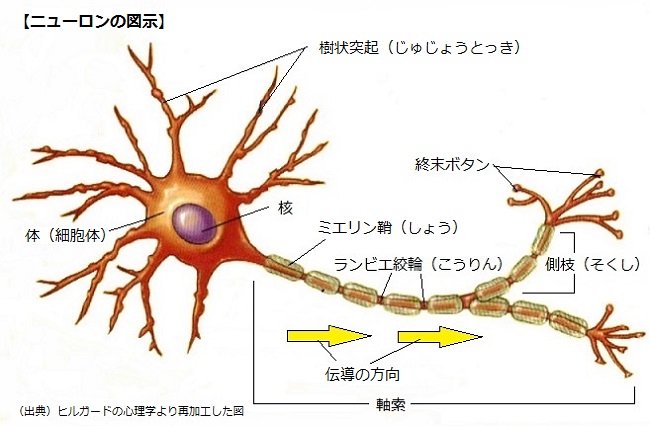
この様にして、ほとんどのニューロンの軸索は、1つのニューロンの樹状突起と細胞体にシナプスを形成し、シナプス後(受けて側の)神経細胞は、複数のシナプス前細胞からの情報を統合する。また、すべてのニューロンはこれらの一般的特徴を持っているが、形や大きさは異なっている。
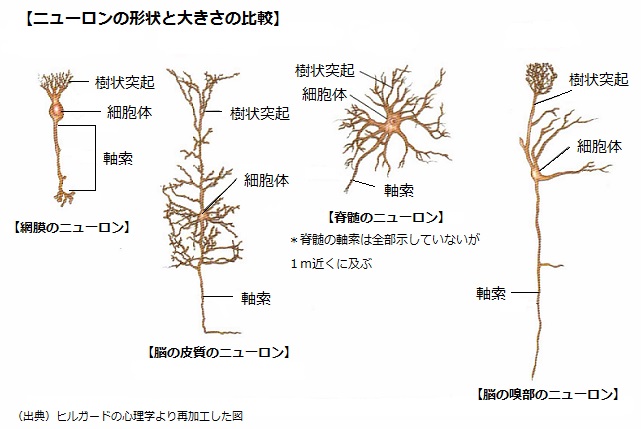
また、ニューロンは一般的機能から3種類に分類される。
1)感覚ニューロン(sensory neuron)
受容器(receptor)からのインパルスを中枢神経へ伝達する。受容器は感覚器官、筋肉、皮膚、関節などにあり、物理的あるいは化学的な変化を感知し、感覚ニューロンを伝わるインパルスに変換する。
2)運動ニューロン(motor neuron)
脳や脊髄から筋肉や内分泌腺などへ、外向きに出て行く信号を運ぶ。
3)介在ニューロン(interneuron)
感覚(求心性)ニューロンと運動(遠心性)ニューロンを接続する。また、介在ニューロンは脳、目のみで見つかっている。
*求心性、遠心性については神経系の基本的な構成を参照。
<神経(nerve)>
ニューロンの軸索が何百、何千からなる細長い束になっていて、たとえば、視神経は信号を目から脳へ運ぶ、また、1つの神経は感覚ニューロンと運動ニューロンの両方の軸索を含む場合もある。ニューロンの細胞体は、一般的に神経系全般で群れを形成している。
<神経核(nucleus)>
脳と脊髄におけるニューロンの細胞体の集まりを神経核(ganglion)と呼び、脳と脊髄以外の細胞体の集まりは神経節と呼ばれる。
<グリア細胞(glial cell)>
ニューロンに加え、神経系には神経以外の細胞も含んでいて、この細胞をグリア細胞(glial cell)と呼ばれる。この細胞の多くは、ニューロンの間にそれを取り囲むようにして散在していて、また、グリア細胞の数はニューロンの9倍あり、脳の容積の半分以上を占めている。
さらに、グリア細胞はニューロンに栄養を与え、老廃物を集め包み込んだりして死んだ細胞や異物を食べ、脳内をきれいに掃除し、ニューロンの信号伝達能力を維持する働きがある。
活動電位((action potential)インパルス)と伝導の仕組み
神経細胞膜の脱分極が一定の臨界値を超える時に、細胞膜に生じる持続時間の短い(1~5ミリ秒)、大きな(60~100mV)脱分極性の膜電位変化で一過性のもの、つまり、細胞膜の一局所が興奮し、そこに発生した活動電位が原因となって隣り合う細胞膜に次々と活動電位が発生し興奮が伝導される時、その伝導される活動電位をインパルスと呼ばれる。
また、インパルスは活動電位あるいはスパイク電位の同義語として用いられる事が多いが、特に神経の軸索を伝導するスパイク電位についての呼び方である。
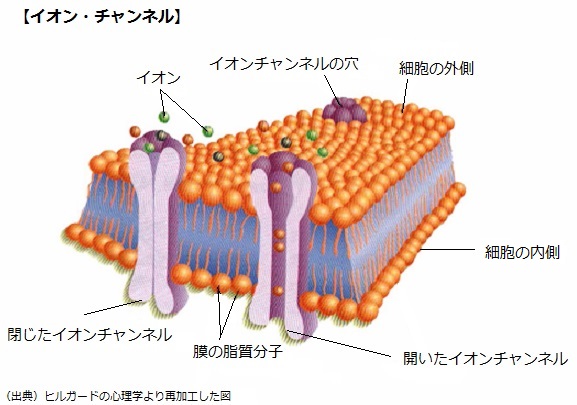
①イオンチャンネル(ion channel)
ドーナツ型のたんぱく質分子で、細胞膜を貫通する穴を形成し、ナトリウムイオン(Na+)、カリウムイオン(K+)、塩素イオン(Cl-)の細胞内、外への流動を調節して、また、それぞれのチャンネルには選択性があり、それが開いている時、一つのイオン(ion)だけが通過できる仕組みになっている。
②静止電位(resting potential)
ほとんどのニューロンでは、静止電位は約-70mv(ミリボルト)である。
ニューロンが活動電位を発生していない静止状態にあるときは、ナトリウムイオン(Na+)を通過させず、細胞外に高く保ち細胞内を低く保つ。電気的には内側と外側を分極させ、内側を外側に比べ、より陰性(マイナス)に保つ、この静止時にある電位を静止電位(resting potential)と呼ぶ。
③興奮閾値((excitation threshold)こうふんいきち)・脱分極(depolarized)
多くのニューロンでは-55mv(静止電位より15mv高い)まで電位が上昇した場合、細胞膜は一過性に不安定になり活動電位が発生する。そのとき、Na+チャンネルが開き、Na+イオンは膜を透過して細胞内に流入する。
相反する荷電は引き受けあうので、陽性に荷電したNa+イオンはさらに流入し、その場の軸索の内側は外側に比べ、+40mv程度上昇し、そして、K+チャンネルが開き、他の陽性に荷電したイオン(K+イオン)は、外側に流出する。
そのことにより、膜の電位を一過性に反転させる脱分極(depolarized)が起こる。
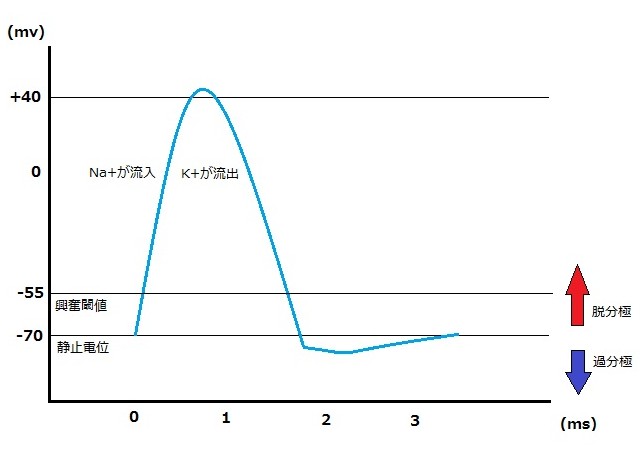
つまり、軸索のランビエ絞輪の部分では細胞膜の内側と外側に電位差(静止電位)があり、その状態(静止状態)が強い刺激により閾値以上に上昇するとNa+に対する細胞膜の透過性が高まり、Na+が細胞膜の外側から内側に流入し、そしてK+に対するイオンチャンネルが開き急激に流出し、そして、静止電位で内部を外部に比べ陰性(マイナス)に分極していた状態が、脱分極によって+40mv程度の活動電位が発生する。
また、この発生した活動電位は、軸索を終末ボタンに向かって隣接した領域を次々と脱分極させ同じ大きさで伝わる。また、インパルスが起きた後の数千分の一秒の間、発生した膜では興奮性が低下するためインパルスが発生せず刺激に反応しないようになっています。これを不応期(refractory period)と呼ばれます。
この不応期によってインパルスは発生した部位から離れた方向に伝わり、反対方向には伝わらず一方方向へ伝わる。
また、この軸索はランビエ絞輪以外はミエリン鞘により絶縁されているため、インパルスはランビエ絞輪間を次々に飛んで伝導され、このような伝導を跳躍伝導(saltatory conduction)と呼ばれる。
インパルスは脱分極が一定の閾値(いきち)を超えたときにだけに発生し、それは「起こる」か「起こらない」かなので、もしインパルスが起こればその大きさは常に同じ大きさで起こる。この法則を神経活動の「全か無かの法則(all-or-none law)」と呼ばれる。
シナプスでのインパルス(活動電位)の伝達
「ニューロンの構造」で説明したように、ニューロン間は直接シナプスの部分で接合していないので、信号はこの隙間を越えて伝達される。
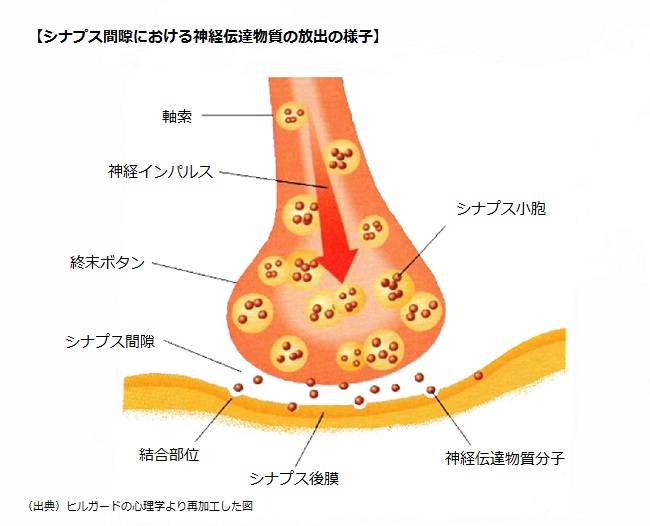
伝達の仕組みは、活動電位が軸索に伝わり終末ボタンに到達すると、終末ボタンに多数ある球状のシナプス小胞が刺激され、シナプス小胞の中には神経伝達物質が含まれており、刺激されたシナプス小胞は、神経伝達物質をシナプス間隙に放出する。
神経伝達物質は、送り手側、すなわちシナプス前ニューロンから、シナプス間隙を拡散していき、受容体に結合する。神経伝達物質と受容体部位は、ジグソーパズルの切片や鍵と鍵穴のようにぴったりと結合する。
この鍵と鍵穴の作用 (lock-and-key action)は、受け手側のイオン・チャンネルの透過性を変化させる。
また、受容体に結合したとき神経伝達物質の中には、興奮性(excitatory)もしくは抑制性(inhibitory)のいずれかの効果を示す。
興奮性の効果により、神経伝達物質がNa+イオンのような陽性に荷電されたイオンを透過させ、それにより受け手側のニューロンが脱分極を起こり、そして、細胞の内側が外側に対して相対的に陽性になることで、後シナプスニューロンは興奮閾値により達しやすく、よりインパルスが発生しやすくなる。のいずれかの効果を示す。のいずれかの効果を示す。
受け取る側の神経細胞のイオンチャンネルの透過性の変化が抑制性となる。また、陽性に荷電したイオン(カリウムのような)が神経細胞から流出たり、陰性に荷電されたイオン(塩素)が流入が引き起こされ、静止電位よりも分極した状態、過分極((hyperpolarized)内側が以前に比べより陰性に荷電される)を起こす。
結果として、興奮閾値に達する確率が低くなり、活動電位が発生する可能性下がります。
主な神経伝達物質の性質と働き
| アセチルコリン | 神経系のいたるところのシナプスに存在し、通常は、興奮性であるが、受け手側ニューロン膜の受容体分子の種類によっては抑制性にもなる。 アセチルコリンは、新しい記憶の形成に重要な役割を果たし、大脳の深部にある大脳基底核や海馬と呼ばれる前脳部分に多く分布している。 また、アルツハイマー型認知症は、アセチルコリンを産出する前脳部位のニューロンが変性する傾向があり、産出が減少すればするほど、記憶障害が重症化するとされる。 |
| ノルエピネフリン | ノルエピネフリンは、主に脳幹部のニューロンにより産出され、これを増加、減少させる薬の効果は、どれも気分の水準を上げたり、下げたりする。 |
| ドーパミン | 中脳の黒質と呼ばれる部分に多く分布しており、脳のある部位でのドーパミン増加は、強い快の感情をもたらす。 ドーパミンが依存形成に果たす役割について研究がされている。 ある脳の部位に、過剰なドーパミンの活動は、統合失調症を起こすかもしれないとされる。 逆に分泌欠如によってパーキンソン病が発症することが明らかにされている。 |
| セロトニン | 脳幹の縫線核に多く分布しており、感情の制御に重要な役割を果たしている。 セロトニンの分泌低下は、抑うつ感情と関連している。 また、睡眠や食欲の調節にも重要な役割があるにで、摂食障害などの治療にも用いられる。 また、分子構造が似ている薬物(LSD)を投与すると、幻覚など気分が高揚することも明らかになっている。 |
| グルタミン酸 | 興奮性の神経伝達物質であるグルタミン酸は、他の神経伝達物質よりも多く脳内に分布している。 三種類以上あるとされ、グルタミン酸受容体の下位類型の一つであるNMDA受容体は、学習や記憶に関係しているとされる。 また、グルタミン酸の神経伝達物質の中断は、統合失調症と関連があるとされている。 |
| GABA | この物質は、主要な抑制性の伝達物質で、実際に脳内のほとんどのシナプスがGABAを使用している。 ピクロトキシンという薬物は、GABAの受容体をふさぎ、けいれんを引き起こす。これは、GABAの抑制の影響なしには、脳による筋肉の動きの抑制ができないということである。 また、不安を改善する薬物は、GABAの活性を強めることで、精神鎮静作用がもたらされる。 |

コメント